Contenu
Le cycle ovarien est constitué de changements répétés dans les ovaires provoqués par les effets des hormones gonadotropes de l'hypophyse. Il se compose de trois périodes différentes, chacune caractérisée par certaines caractéristiques. Un autre nom pour ce concept est le cycle ovarien, ce qui signifie son lien avec l'ovulation.
Physiologie du cycle ovarien
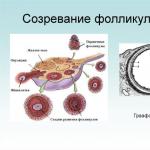
Pour mieux comprendre l'essence du cycle ovarien, il est nécessaire de comprendre plus en détail les changements dans le corps féminin provoqués par les hormones. Ces hormones sont sécrétées par l’hypophyse antérieure et sont appelées hormones folliculo-stimulantes. Le début de la période ovarienne chez une femme adulte commence par le développement de plusieurs follicules primaires. Ils contiennent des ovocytes de premier ordre – la forme immature des futurs ovules.
L’idée est de passer par trois étapes :
- folliculaire;
- ovulation;
- lutéal
La dernière étape est également appelée étape progestérone en raison de la formation du corps jaune, son apparition est provoquée par la progestérone. Un cycle menstruel normal dure 28 jours, donc la première et la dernière étape le divisent en deux. Avec une durée de cycle de 28 jours, 14 jours sont alloués aux phases folliculaire et lutéale. Entre les deux se trouve la phase ovulatoire.
Important! Le cycle ovarien remplit deux fonctions importantes : il favorise la production d'ovules et libère cycliquement des hormones sexuelles. Si une femme est en bonne santé et ne présente aucune inflammation ou pathologie des organes génitaux, cette période survient mensuellement sans interruption, confirmant la fonction reproductrice du corps.
Phases du cycle
Le cycle menstruel est divisé en deux catégories : les cycles ovariens et utérins. Cette dernière se divise également en 4 étapes : desquamation, régénération, prolifération, sécrétion. Chacune de ces étapes est responsable de la production de certaines hormones nécessaires au bon fonctionnement du corps féminin. Parallèlement, au moment de franchir chaque étape, un certain nombre de changements sont observés. Pour une compréhension générale de la cyclicité, il est recommandé de considérer chaque période séparément.
Phase folliculaire (menstruelle)
Le stade folliculaire vient en premier. Il est responsable du développement du follicule dans les ovaires, après quoi l'ovule est libéré. Normalement, ce processus se produit au début du cycle menstruel, souvent au cours des 9 à 14 premiers jours. Si le cycle menstruel dure 21 jours, le follicule devrait alors mûrir en 10 à 11 jours.

Initialement, le follicule contient un ovule au stade de développement, entouré d'une couche épithéliale. La coquille est constituée de tissu conjonctif. Il y a plusieurs caractéristiques de cette étape :
- la croissance se produit sous l'influence d'hormones folliculo-stimulantes ou FSH ;
- un seul des follicules formés mûrit, à partir duquel l'ovule est ensuite libéré ;
- Les petits éléments non mûrs se dissolvent avec le temps.
Le liquide à l’intérieur est rempli d’œstrogènes, qui participent à tous les processus du système reproducteur de la femme. Lorsque le follicule atteint sa pleine maturité, sa taille augmente, ce qui provoque souvent une gêne chez la femme. Lors d'un examen par un gynécologue, le médecin peut facilement confirmer sa maturation. Vous pouvez en apprendre davantage sur la folliculogenèse à partir de la vidéo :
Phase ovulatoire (proliférative)
Au stade de l'ovulation, l'élément mature se rompt. Ce moment est caractérisé par le fait que l'œuf doit être mature et apte à la fécondation. En sortant du follicule, il pénètre dans l'espace abdominal avec le liquide folliculaire, après quoi il est transporté dans la trompe de Fallope.

Avant la rupture, le tissu s’amincit et l’accumulation de liquide folliculaire favorise une rupture rapide. Il existe une opinion selon laquelle il existe un lien entre les ovaires et l'hypophyse : en raison de l'effet sur les centres nerveux du corps, un mécanisme réflexe de rupture se produit sous l'influence d'une irritation.
Important! Dès la naissance, le corps d’une femme contient environ 300 à 400 000 ovules. Pendant la puberté, la première ovulation a lieu et avant la ménopause, la fonction ovulatoire du corps prend fin.
La phase de prolifération est extrêmement importante pour les femmes souhaitant devenir enceintes. Le moment où un œuf mature est libéré est facile à calculer. Habituellement, c'est 3 jours avant l'ovulation, l'ovulation elle-même et 2 jours après.
Phase lutéale (sécrétoire)
Cette étape est également appelée phase du corps jaune. À ce stade, les événements suivants se produisent dans le corps :
- La phase de sécrétion commence pendant l'ovulation et dure jusqu'à la prochaine menstruation.
- Les cellules du follicule, qui se rompent pendant la phase d'ovulation, accumulent des pigments et des graisses. C'est pourquoi il devient jaune et est appelé corps jaune.
- Le corps jaune produit des androgènes, des œstrogènes et des progestérones - les principales hormones sexuelles nécessaires à la grossesse et au fonctionnement normal du corps féminin.
- Si la grossesse ne se produit pas, la synthèse de substances par le corps jaune s'arrête et les règles surviennent.

La durée normale de la période lutéale est de 12 à 16 jours. Ce chiffre dépend de la durée du cycle menstruel.
Troubles possibles du cycle ovarien
Durant toute la période, certaines perturbations peuvent survenir. Par exemple, on sait que dans la première phase du cycle ovarien, une hormone est libérée qui stimule la croissance des follicules. Pour certaines raisons, ce type d'hormone peut ne pas être libéré, ce qui est associé à des troubles :
- faible progestérone dans la phase lutéale causée par une hyperprolactinémie, des pathologies des glandes surrénales, des maladies de la glande thyroïde ;
- taux élevé de progestérone dans la phase lutéale, provoqué par une cystose du corps jaune, un dysfonctionnement rénal, un échec menstruel ;
- maturation prolongée de l'œuf;
- incapacité à ovuler causée par la grossesse, l'allaitement, l'adolescence ou la prise de pilules.

Les causes des troubles de l'ovulation sont également des déséquilibres hormonaux, associés à des pathologies du système endocrinien. Une autre cause de dysfonctionnement du cycle ovarien est l’excès ou l’insuffisance pondérale.
Conclusion
Normalement, une femme en bonne santé a un cycle ovarien tous les mois. Il dit que le corps est capable de se reproduire et que lorsqu'un ovule mature est fécondé, une grossesse se produit. Si une femme ne peut pas tomber enceinte pendant une longue période, il existe des violations qui méritent également d'être recherchées dans le développement de chaque étape.
Le cycle menstruel peut être divisé en deux types : ovarien et utérin. Le cycle ovarien est une séquence de processus naturels de développement de l'ovule et de croissance du corps jaune à partir des parois d'un follicule éclaté, qui durent jusqu'à 27 jours et se répètent périodiquement chez une femme en bonne santé.
Le cycle ovarien complet est divisé en trois phases :
- Folliculaire – pendant lequel les follicules grandissent et se développent.
- La phase ovulatoire est la libération d'un ovule mature par le follicule.
- Lutéal - la formation d'un corps jaune sur le site d'un follicule éclaté.
Phase folliculaire du cycle

Le petit follicule contient l'ovule au stade initial de développement et la couche épithéliale qui l'entoure. La couche externe du follicule est constituée de tissu conjonctif. La première phase du cycle ovarien et la croissance des follicules se produisent sous l'influence de la FSH ; un seul d'entre eux arrive à maturité. Les petits follicules se dissolvent avec le temps.
Les follicules mûrissent au début du cycle ovarien et ce processus dure de 9 à 13 jours. Chaque composant du follicule subit des changements spectaculaires au cours de cette phase. La taille de l’ovule sera multipliée par cinq et son protoplasme et sa membrane se prépareront à quitter le follicule. Le développement se terminera après une double division. Division initiale (maturation) - la formation de deux cellules nerveuses. La cellule principale continuera à se diviser et la seconde se dissoudra. À la fin de la division de réduction (deuxième), l'œuf est complètement mature et peut devenir un zygote.
Important! L'épithélium du follicule au cours du développement subira une prolifération - il deviendra multicouche et se transformera en une membrane folliculaire granulaire.
À l'avenir, sur le site de la plus grande accumulation de cellules, des vides au remplissage uniforme apparaîtront. Bientôt, ils fusionneront, formant une cavité continue avec du liquide folliculaire. Au cours du processus d'accumulation de liquide, la coque granulaire se dilate d'un côté et se rétrécit de l'autre. Les cellules situées plus près de l’œuf sont appelées couronne radiée. Par la suite, l'ovule est situé sur les cellules accumulées près de la paroi - le monticule ovipare. Lorsque le follicule est complètement mature, l’ovule change de position et prend sa place dans le liquide folliculaire.
Lors de la croissance du follicule, sa coquille forme deux couches :
- Interne - contient des capillaires et des éléments cellulaires.
- Structure externe - fibreuse avec de gros vaisseaux.
Le liquide à l'intérieur du follicule et la couche granulaire sont remplis d'hormone œstrogène, qui participe au fonctionnement de l'ensemble du système reproducteur féminin. Vers la fin de la maturation, le follicule va augmenter en taille.
Phase ovulatoire

La période ovulatoire est la période pendant laquelle un ovule mature quitte le sac folliculaire. La cellule mature quittera le follicule avec la couronne radiée et le liquide environnant et sera transportée dans la trompe de Fallope. La fécondation a lieu à l'intérieur du tube. Si le sperme ne pénètre pas dans l’ovule, il perdra rapidement sa viabilité (jusqu’à 24 heures) et se dissoudra. La rupture des parois folliculaires ne provoque pas de saignement grave ; une petite quantité de sang peut s'écouler des capillaires endommagés.
L'ovulation se produit en raison des effets combinés des gonadotrophines et des œstrogènes. La synthèse intensive de LH a une grande influence. Pour réussir à rompre les parois du sac folliculaire, le liquide à l'intérieur augmente de volume et les parois de l'ovaire deviennent plus fines. Si le cycle menstruel dure 27 jours, l'ovulation aura lieu environ avant le 14ème jour. La période avant et après l’ovulation est la plus propice à la fécondation.
Phase lutéale ou période de croissance du corps jaune
Le corps jaune est la composante temporaire de l’ovaire, caractérisée par une vascularisation et une stéroïdogenèse accrues. Le corps jaune remplit ses fonctions pendant 13 jours et se dissout par conséquent.
Le principal indicateur de la phase lutéale est la production intense de progestérone. Pendant cette période, la température d’une femme peut augmenter en raison de l’effet de la progestérone sur la thermorégulation. La production de progestérone est maximale (24 mg par jour) le huitième jour après une sécrétion accrue de LH. La teneur en 17-hydroxyprogestérone, estrone et estradiol augmente également. Dans le même temps, les niveaux de LH et de FSH diminueront.
Fonction endocrinienne de la TV

- Production de progestérone. L'intensité de la production de progestérone dépend du fonctionnement des cellules lutéales. Cela ne sera suffisant que si...
- Au début du cycle, l'effet de la FSH sur les cellules de la granulosa était intense, de nombreux récepteurs de FSH et de LH se formaient et la production d'œstradiol était dans les limites normales.
- Dans la première phase du cycle, la sécrétion de FSH est supprimée sélectivement et une petite quantité d'œstrogène est libérée. La phase mi-lutéale est caractérisée par des taux de progestérone plus faibles et une diminution du nombre de cellules lutéales. Il en résulte un raccourcissement de la phase lutéale et une faible réponse cellulaire à l'hCG.
- Les lipoprotéines de basse densité qui traversent le lit vasculaire de l'AT jusqu'aux cellules lutéinisées doivent se lier aux récepteurs de ces cellules et sont capables de déclencher la production de progestérone. Ils sont une source de cholestérol, qui contribue à son tour au fonctionnement normal du corps jaune. Si l'apport sanguin est altéré et que les lipoprotéines diminuent, la phase lutéale sera défectueuse.
- La sécrétion de LH se situe dans les limites normales. Si la sécrétion diminue, cela affectera les niveaux de progestérone, ce qui entraînera l'apparition prématurée des règles.
- Synthèse de relaxine. La relaxine est une hormone de type peptidique qui reflète l'appareil ligamentaire de l'articulation interpubienne et contrôle le fonctionnement du myomètre. Lorsqu'elle est stimulée par l'hCG, la production de relaxine augmentera au milieu de la troisième phase. Cette hormone vous permet de maintenir une grossesse dès les premiers stades.
- Peptides de type vasiopressino et ocytocine. Les principales fonctions de ces peptides sont parkrine et endocrinienne. Ces fonctions contrôlent le microenvironnement ovarien, sa modification entraînera l'ovulation. L'ocytocine, associée à la prostaglandine, favorise la libération de l'ovule du follicule. La vasopressine participe au contrôle de la microcirculation dans l'ovaire et possède également des propriétés vasoconstrictrices.
Destruction du corps jaune

VT passe par 4 phases de développement :
- Prolifération;
- Vascularisation ;
- Floraison;
- Destruction.
Lorsqu’il est détruit, le corps jaune est appelé menstruel. Étant donné que la vasopressine et l'ocytocine sont en concentrations élevées dans la TV, elles affectent ses fonctions. Au cours de la régression, la diminution de la stéroïdogenèse dans les cellules lutéales est influencée par la fréquence des impulsions de sécrétion de LH.
Bientôt, le VT se transformera en une formation appelée corps blanc. C'est une cicatrice qui disparaîtra avec le temps.
Si une femme tombe enceinte, la TV ne s'effondrera pas, mais continuera à fonctionner jusqu'à environ 18 semaines, pendant la formation du placenta. Après cela, le corps jaune se dissoudra.
Transition vers un nouveau cycle

Le développement des premiers follicules ne commencera qu’après la destruction du corps jaune. La TV disparaît quelques jours avant le début des règles. Parallèlement à la lutéolyse, les taux de progestérone et d'estradiol diminuent. À son tour, la sécrétion de LH et de FSH va augmenter. Le cycle ovarien se répétera. La synthèse des hormones stéroïdes et la folliculogenèse se produisent continuellement de la période prénatale à la période postménopausique.
Cycle utérin

Cette partie du cycle menstruel est divisée en quatre phases : desquamation, régénération, prolifération, sécrétion.
La durée normale de prolifération est de 12 à 15 jours. Les changements qui se produisent au cours de cette période au niveau de l'endomètre sont provoqués par une production importante d'œstrogènes. Dans les premiers jours de la phase de prolifération, la couche fonctionnelle ne commence pas encore à se diviser en zones ; la surface est recouverte d'un épithélium cylindrique de forme cubique, l'endomètre est aminci. Des cryptes glandulaires de forme ronde ou ovale sont visibles en coupe transversale. Ils ressemblent à des tubes droits ou légèrement courbés avec une fine lumière.
Au milieu de la prolifération, l'activité de la phosphatase alcaline dans l'endomètre augmente. Un relâchement et un gonflement apparaissent au niveau de l'étrier. Par rapport au début du cycle, le nombre de mitoses augmente. Le cytoplasme et les noyaux des cellules stromales sont mieux visualisés. Le stroma, comme auparavant, a des parois minces.
A la fin de cette phase (au 13ème jour du cycle), la division en zones n'a pas encore eu lieu, mais on peut noter un compactage de la couche fonctionnelle. Les structures glandulaires changent de forme en forme de bouclier et sont plus proches les unes des autres.
La phase de sécrétion (jusqu'à 15 jours) est inextricablement liée à la fonction endocrinienne du corps jaune et à la production de progestérone. Si la durée de la phase chez une femme en âge de procréer augmente ou diminue de trois jours. C'est le premier signe de perturbations du système reproducteur. Des changements de durée se produisent pendant la ménopause.
Note: Le début de cette phase est caractérisé par la croissance de l'endomètre. Le principal signe du début de la phase de sécrétion est l'apparition de vacuoles sous-nucléaires dans les glandes épithéliales. Le milieu de la phase de sécrétion survient le 22ème jour du cycle. Des changements importants au cours de cette période se produisent dans l'endomètre sous l'influence des hormones produites par le corps jaune. La couche fonctionnelle est épaissie comme auparavant. Il existe une division en couches profondes et superficielles. Dans la couche superficielle, la tortuosité des cryptes glandulaires est insignifiante et il existe un grand nombre de cellules stromales. L'épithélium de cette couche est visualisé sous la forme d'un prisme, non sécrétant.
La fin de la phase de sécrétion survient entre 23 et 26 jours si la conception n'a pas eu lieu. Des processus régressifs se produisent dans l'endomètre, provoqués par la mort du corps jaune et une diminution de la quantité de progestérone.
Au cours de la dernière phase du saignement, une desquamation et une régénération ultérieure se produisent.
Vidéo : Processus pendant le cycle menstruel
Le principal moyen de neutraliser l'ammoniac.
4.Coenzyme : concept, classification, exemples.
Répondre:
Page 210
2) Cycle ovarien :
1- phase folliculaire : développement folliculaire, sécrétion d'œstrogènes et ovulation
2-phase lutéale : le corps jaune fonctionne, la progestérone est sécrétée
3 phases d'involution du corps jaune : la sécrétion d'oestrogène et de progestérone s'arrête
phase folliculaire: La FSH provoque la maturation des follicules et la formation d'œstrogènes. La libération d'œstrogènes dans le sang inhibe la sécrétion de FSH et stimule la formation de LH, qui assure l'ovulation et la production de progestérone, le passage à la phase suivante.
phase lutéale: le corps jaune se forme, qui produit de la progestérone qui, entrant dans le sang, inhibe la sécrétion de LH et stimule la libération de prolactine. La prolactine soutient la production de progestérone et stimule le développement des glandes mammaires. Si l’ovule n’est pas fécondé ou implanté, la transition vers la phase 3 commence. En cas de fécondation, une grossesse se produit.
phase d'involution du corps jaune : le corps jaune subit un développement inverse, la production de progestérone diminue progressivement. De faibles niveaux d’œstrogène et de progestérone dans le sang entraînent la réactivation de la production de follibérine et de FSH et, par conséquent, la phase folliculaire commence.
Les phases du cycle ovarien correspondent à certains changements dans l'utérus provoqués par les hormones sexuelles - les phases utérines.
Cycle utérin :
1- phase proliférative : les œstrogènes libérés lors de la maturation du follicule agissent sur l'endomètre, provoquant une prolifération de l'épithélium utérin et une augmentation de l'activité contractile du myomètre.
2- phase de sécrétion : l'endomètre, préparé par les œstrogènes, sous l'influence de la progestérone, sécrète du mucus, celui-ci est nécessaire à l'implantation de l'ovule ;
3ème phase menstruelle : la production de progestérone se poursuit, ce qui inhibe la production de LH. Une diminution de la LH provoque un rejet de la muqueuse et des saignements.
3) L'ammoniac est neutralisé des manières suivantes :
a) amination réductrice (insignifiante, bien qu'elle assure la formation de certains acides aminés)
b) la formation d'amides d'acide aspartique et glutamique - asparagine et glutamine. Ce processus se produit dans les tissus nerveux, musculaires et rénaux ; les catalyseurs sont l'asparagine synthétase et la glutamine synthétase.
c) la formation de sels d'ammonium se produit dans le tissu rénal, où l'ammoniac est délivré sous forme d'asparagine et de glutamine. Ici, ils sont hydrolysés pour former de l'aspartate et du glutamate, et de l'ammoniac est libéré. L'ammoniac est neutralisé par la formation de sels d'ammonium, qui sont éliminés dans les urines.
d) la synthèse de l'urée - principal moyen de neutralisation et d'élimination de l'ammoniac - s'effectue dans le foie. Cela se produit dans plusieurs réactions :
1 – synthèse du phosphate de carbamoyle ; enzyme – carbamoylphosphosynthétase.
2 - le phosphate de carbamoyle interagit avec l'ornithine pour former de la citrulline ; catalyseur – ornithine carbamoyl phosphate transférase.
3 – la citrulline interagit avec l’aspartate pour former le succinate d’arginine.
4 – le succinate d’arginine est décomposé en fumarate et arginine.
5 – l'arginine, sous l'action de l'arginase, est décomposée hydrolytiquement en urée et ornithine.
L'urée est un composé inoffensif, sa synthèse se produit dans le foie, dont le dysfonctionnement entraîne un ralentissement du processus, une diminution de la teneur en urée dans le sang et une diminution de l'excrétion dans les urines.
4) Coenzymes – Ce sont des substances nécessaires à l’activité de certaines enzymes. Ils participent directement à la réaction chimique catalysée par l'enzyme.
Classification:
a) inorganique (ions métalliques, certains anions)
b) biologique
Ions métalliques – ions calcium, magnésium, potassium, zinc, fer. Ils interviennent dans : la stabilisation de la structure tertiaire ou quaternaire, la fixation du substrat ou la catalyse.
Il existe des coenzymes nature des nucléotides, coenzymes tétrapyrrole et coenzymes - dérivés de vitamines.
Coenzymes – nucléotides – dans le cadre des transférases, ils participent au transfert du phosphate, du pyrophosphate, de l'adénylate et à la transformation des sucres.
Coenzymes tétrapyrrole identique à l'hème dans l'hémoglobine ; participer au transport des électrons dans le cadre des cytochromes et de la peroxydase.
Coenzymes - vitamines participer à diverses réactions métaboliques chimiques. Par exemple, la forme coenzyme de la vitamine B1 (thiamine), le diphosphate de thiamine, catalyse la réaction de décarboxylation.
Cycle menstruel.
Chez l’homme, la formation et la libération des gamètes sont un processus continu qui commence dès le début de la puberté et se poursuit tout au long de la vie. Chez la femme, il s'agit d'un processus cyclique qui se répète environ tous les 28 jours et est associé à des changements dans la structure et les fonctions de l'ensemble du système reproducteur. Ce processus s’appelle le cycle menstruel et peut être divisé en quatre étapes. Les événements qui surviennent au cours du cycle menstruel impliquent les ovaires (cycle ovarien) et l'utérus (cycle utérin) et sont régulés par les hormones ovariennes, dont la sécrétion est à son tour régulée par les gonadotrophines hypophysaires.
Cycle ovarien.
Chez la femme adulte, le cycle ovarien débute par le développement de plusieurs follicules primaires (contenant des ovocytes de premier ordre) sous l'influence de l'hormone folliculo-stimulante (FSH) sécrétée par l'hypophyse antérieure. Parmi ces follicules, un seul continue de croître, tandis que les autres sont détruits à la suite d'un processus dégénératif (atrésie folliculaire). Les cellules de la membrane de la granulosa du follicule en croissance prolifèrent, formant une couche fibreuse externe de plusieurs cellules d'épaisseur, appelée thèque extema, et une couche interne riche en vaisseaux sanguins, thèque interne. Les cellules de la granulosa sécrètent du liquide folliculaire qui s'accumule dans la cavité folliculaire. L'hormone lutéinisante (LH), sécrétée par l'hypophyse, stimule les cellules, les amenant à produire des stéroïdes, principalement de l'estradiol. L'augmentation des taux d'estradiol au cours de la phase folliculaire agit sur l'hypophyse selon le principe du feedback négatif, provoquant une diminution des taux de FSH dans le sang (jours 4 à 11) ; Les niveaux de LH restent inchangés. Les niveaux d'œstrogènes culminent environ trois jours avant l'ovulation et agissent à ce moment-là sur l'hypophyse selon le principe de la rétroaction positive, stimulant la libération de FSH et de LH. On pense que la FSH est nécessaire pour stimuler la croissance folliculaire, mais le développement folliculaire ultérieur est principalement contrôlé par la LH. Les cellules de la granulosa sont situées à la périphérie, l'œuf est déplacé d'un côté du follicule, mais est toujours entouré d'une couche de cellules de la granulosa. Le follicule mature, appelé vésicule de Graaf, atteint environ 1 cm de diamètre et dépasse de la surface de l'ovaire sous la forme d'un tubercule. Le mécanisme exact de l’ovulation est inconnu, mais on pense que la LH, la FSH et les prostaglandines sont impliquées.
Pendant l'ovulation, un ovocyte de second ordre se sépare de la paroi du follicule rompu, sort dans la cavité abdominale et pénètre dans la trompe de Fallope.
Généralement, chaque mois, un seul ovocyte est libéré par l'un des ovaires, de sorte que l'ovulation se produit alternativement dans l'un ou l'autre ovaire. Un ovocyte ovulé est une cellule dont le noyau est en métaphase 1 de la méiose ; elle est entourée d'une couche de cellules appelée zone pellucide et d'une couche de cellules de la granulosa (corona radiata), qui protège l'ovocyte jusqu'à la fécondation. Après l'ovulation, le niveau de LH chute au niveau caractéristique de la phase folliculaire et sous l'influence d'une autre gonadotrophine - la prolactine - les cellules du follicule rompu changent, formant le corps jaune. Le corps jaune commence à sécréter une autre hormone féminine – la progestérone – et de petites quantités d’œstrogène. Ces deux hormones maintiennent la structure normale de l'endomètre tapissant l'utérus et inhibent la libération de FSH et de LH, agissant sur le principe du feedback négatif sur l'hypothalamus. Si la fécondation ne se produit pas, alors sous l'influence de facteurs qui seront décrits plus loin, le corps jaune subit une involution et seule une petite cicatrice reste - le corps albicans ; cela s'accompagne d'une diminution des niveaux de progestérone et d'œstrogène, à la suite de laquelle l'inhibition de la sécrétion de FSH cesse, son niveau augmente et un nouveau cycle de développement folliculaire commence.
Cycle menstruel- des changements cycliques dans le corps d'une femme, dont la manifestation externe est la menstruation.
Les modifications cycliques des ovaires - le cycle ovarien - sont divisées en phases folliculaires et lutéales, et les modifications de l'endomètre - le cycle utérin - en phases prolifératives et sécrétoires. À la suite du rejet de la couche fonctionnelle de l'endomètre, la menstruation survient. La ménarche est la première menstruation. Il est généralement observé entre 10 et 12 ans, tandis qu'un cycle régulier s'établit généralement après 1 à 1,5 ans. Le cycle moyen est de 28 jours, la normale est de 21 à 35 jours. Le premier jour des règles correspond au premier jour du cycle menstruel. La durée des menstruations est de 2 à 7 jours (en moyenne 4 à 5 jours), la perte de sang est de 50 à 150 ml (en moyenne 70 à 100 ml).
Le cycle menstruel est déterminé par le travail conjugué de cinq maillons de la chaîne neurohumorale (cortex cérébral, hypothalamus, hypophyse, ovaires, utérus).
Les principaux produits de sécrétion de l'hypothalamus sont des facteurs de libération hypophysaires. L'hormone de libération des gonadotrophines (GnRH) contrôle la sécrétion des gonadotrophines hypophysaires, des hormones lutéinisantes (LH) et des hormones folliculo-stimulantes (FSH) (Fig. 4).
La GnRH est la seule hormone qui régule la sécrétion pulsatile de deux hormones hypophysaires. La perfusion chronique de GnRH ne stimule pas la sécrétion de gonadotrophines. Le mode pulsatile de sécrétion de GnRH (Fig. 5) est nécessaire en raison de la très courte période de puluralyse RG - environ 2 à 4 minutes. Au cours du cycle menstruel, la fréquence et l'amplitude des pulsations de la GnRH changent : en phase folliculaire, elles sont élevées et en phase lutéale, elles diminuent.
Riz. 4. Régulation du cycle menstruel
 Riz. 5. Modèle de sécrétion de GnRH
Riz. 5. Modèle de sécrétion de GnRH
La fin de chaque cycle menstruel et le début du suivant sont caractérisés par de faibles niveaux de stéroïdes sexuels : progestérone et œstrogènes.
Avec l'arrêt de la fonction du corps jaune, la production de FSH et de LH augmente. Les cellules de la granulosa interagissent avec la FSH et les cellules de la couche thécale interne interagissent avec la LH. Chaque cycle menstruel(Fig. 6) de 3 à 30 follicules primordiaux, sous l'influence de la FSH, entrent en phase de croissance et sécrètent des œstrogènes dont le taux augmente progressivement au cours de la 1ère - phase folliculaire cycle menstruel.
Lors de la croissance des follicules secondaires (au 8ème jour du cycle menstruel), dominant follicule se transformant en follicule tertiaire(préovulatoire, ci-après graphiques bulle, jusqu'à 2-3 cm de diamètre).
La synthèse des œstrogènes s'effectue de deux manières. La première voie implique l'aromatisation enzymatique des androgènes en œstrogènes par les cellules de la granulosa. La deuxième voie est associée à la synthèse d'œstrogènes dans les cellules thécales aux derniers stades de la phase antrale. Ainsi, au milieu de la phase folliculaire, le taux d'œstrogènes et d'androgènes folliculaires augmente, ce qui s'accompagne d'une diminution de la concentration de FSH (feedback négatif).
Parallèlement, les œstrogènes stimulent la sécrétion de LH tout au long de la période folliculaire.
La FSH stimule :
Phase de croissance des follicules primordiaux ;
Transport de liquide dans la cavité folliculaire ;
Expression des récepteurs de la LH et de l'acide prolactique sur les cellules de la granulosa ;
Activité aromatase. La LH stimule :
Production par les cellules folliculaires de protéines de faible poids moléculaire qui neutralisent le facteur qui supprime la méiose ;
Division méiotique de l'ovocyte et passage au stade de 2ème ordre - ensemble haploïde ;
Synthèse des androgènes - androstènedione et testostérone - dans les cellules la CA;
Synthèse de progestérone (lutéinisation) dans les cellules folliculaires ;
Synthèse des prostaglandines dans les cellules folliculaires ;
Induction de l'ovulation.
En phase préovulatoire, sur les cellules de la granulosa du follicule, la FSH induit le développement de récepteurs à la LH et à la prolactine. Ainsi, à la fin de la phase préovulatoire, la teneur en FSH et LH augmente et les cellules folliculaires deviennent insensibles aux effets des œstrogènes et des androgènes. Des concentrations élevées d'œstrogène stimulent la libération de LH et la rupture de la paroi de la vésicule de Graaf (follicule tertiaire) - c'est-à-dire ovulation, survenant 10 à 12 heures après le pic des taux de LH. Ensuite, l'œuf est libéré dans la cavité abdominale et commence phase lutéale du cycle.
Œstrogènes :
Stimuler la prolifération des cellules folliculaires ;
Stimuler l'expression des récepteurs FSH ;
Ils participent (avec la FSH) à la formation des récepteurs de LH dans les cellules folliculaires ;
Augmenter la sécrétion de LH ; avec une teneur élevée en œstrogènes, la GnRH stimule les cellules qui synthétisent la LH ;
Supprimer la sécrétion de FSH ; lorsque les niveaux d’œstrogènes sont faibles, la GnRH stimule les cellules qui synthétisent la FSH.
Androgènes :
Inhiber l'expression des récepteurs FSH sur les cellules de la granulosa ;
Inhibe l'activité de l'aromatase.
Les capillaires résultants se développent rapidement dans la cavité folliculaire, les cellules de la granulosa subissent une lutéinisation sous l'influence de la LH, ce qui conduit à la formation du corps jaune.
Les niveaux d'œstrogènes commencent à diminuer à partir de la fin de la phase préovulatoire dans un contexte de concentrations élevées de FSH et de LH, continuent de baisser au début de la phase lutéale et augmentent à nouveau en raison de la sécrétion du corps jaune.
Corps jaune(corps jaune) est une glande endocrine transitoire qui fonctionne pendant 8 à 14 jours, quelle que soit la durée du cycle menstruel, et synthétise la progestérone, les œstrogènes (principalement le 17b-estradiol) et la prolactine. Les niveaux de progestérone augmentent progressivement après l'ovulation et atteignent leur maximum 8 à 9 jours après l'ovulation, soit approximativement au moment de l'implantation. L'effet thermogénique de la progestérone entraîne une augmentation de la température corporelle d'au moins 0,33°C (l'effet dure jusqu'à la fin de la phase lutéale).
Progestérone :
Prépare l'endomètre à la nidation ;
Détend les fibres du myomètre ;
A un effet natriurétique, stimulant la sécrétion d'aldostérone ;
La progestérone placentaire est métabolisée dans le cortex surrénalien fœtal et les testicules en tant que précurseur des corticostéroïdes et de la testostérone, respectivement.
Ainsi, la phase lutéale est caractérisée par des concentrations accrues de progestérone et de prolactine et de faibles taux de FSH et de LH.
À mesure que la fonction du corps jaune régresse, les concentrations de stéroïdes sexuels diminuent et un nouveau cycle menstruel commence.
En plus des hormones énumérées, le corps jaune puis le placenta produisent de la relaxine. Il inhibe l'activité contractile du myomètre en activant l'action de la progestérone et en augmentant le niveau d'AMPc tant dans les cellules musculaires lisses du myomètre que dans les chondrocytes de la symphyse pubienne, provoquant son ramollissement.
Corps blanc- cicatrice du tissu conjonctif à la place de la fonction achevée et corps jaune dégénéré.
CYCLE OVARIAL
Le nombre d'ovogonies dans un embryon femelle au milieu du développement intra-utérin atteint 5 à 7 millions, cependant, une partie importante des ovocytes subit une atrésie (Fig. 7), associée à une faible production d'hormones gonadotropes. Les ovaires d’une fille nouveau-née contiennent déjà 1 à 2 millions d’ovocytes ; à la puberté, il y en a de 100 à 400 000. Pendant la période de reproduction, 98 % des follicules primordiaux meurent, environ 2 % atteignent le stade de follicules primaires et secondaires, mais pas plus de 400 à 500 ovulent. Tous les follicules qui ont commencé à se développer mais n’ont pas atteint le stade de l’ovulation subissent une atrésie.
Ainsi, la durée de vie d’un ovocyte peut atteindre 40 à 50 ans. C’est pourquoi le risque de malformations génétiques fœtales augmente considérablement avec l’âge de la mère.
 Riz. 7. Modèle de sécrétion de GnRH
Riz. 7. Modèle de sécrétion de GnRH
STRUCTURE FOLLICULAIRE
Follicule primordial recouvert d'une seule couche de cellules folliculaires (granulosa) et entouré d'une membrane basale. Les cellules de la granulosa entourant l'ovocyte (« corona radiata ») sécrètent un substrat glycoprotéique, qui forme une zone transparente - zone pellucide- entre les cellules de l'ovocyte et de la granulosa. À la surface de la zone pellucide se trouvent des récepteurs spécifiques à l'espèce qui interagissent uniquement avec les spermatozoïdes allogéniques ; la pénétration de la zone par un spermatozoïde conduit au développement d'une « réaction zonale » qui empêche la polyspermie.
Les cellules du stroma ovarien forment une couche de cellules fusiformes - la thèque. Les androgènes, qui assurent le développement des follicules, sont produits exclusivement par les cellules thécales. En raison de la prolifération, ces dernières sont divisées en deux couches : la couche interne, qui a une structure glandulaire, et la couche externe, le tissu conjonctif. Entre eux s'accumule le liquide folliculaire, contenant le transsudat sérique et la sécrétion de mucopolysaccharides des cellules de la granulosa. L'accumulation de liquide donne au follicule l'apparence d'une vésicule et un tel follicule est appelé
est secondaire ou antrale (Fig. 8). L'ovocyte qui s'y trouve n'a pas encore subi la deuxième division méiotique.
Le follicule primordial est recouvert d'une seule couche de cellules folliculaires (granulosa) et entouré d'une membrane basale.
 Riz. 8. Croissance folliculaire
Riz. 8. Croissance folliculaire
 Riz. 9. Follicule tertiaire (« vésicule de Graaf »)
Riz. 9. Follicule tertiaire (« vésicule de Graaf »)
Le développement de l'ovocyte se poursuit jusqu'à la fécondation, et la transformation d'un ovocyte de 1er ordre en un ovocyte de 2ème ordre, qui possède déjà un ensemble haploïde de chromosomes, se produit soit immédiatement avant l'ovulation, soit dans la trompe de Fallope.
Au cours de chaque cycle ovarien, 15 à 20 follicules se développent dans les ovaires. Certains d'entre eux atteignent de grandes tailles (jusqu'à 8 mm) au milieu du cycle, mais un seul follicule devient un follicule tertiaire mature d'un diamètre de 2 à 3 cm (« vésicule de Graaf », Fig. 9).
CYCLE UTÉRIN
Les changements dans les niveaux hormonaux affectent directement l'état de l'endomètre, de la membrane muqueuse des trompes de Fallope, du canal cervical et du vagin. La muqueuse utérine subit des modifications cycliques (phases prolifératives, sécrétoires et menstruelles). L'endomètre est divisé en couches fonctionnelles (disparaissant pendant la menstruation) et basales (préservées pendant la menstruation).
Phase proliférative- la première moitié du cycle - dure du premier jour des règles jusqu'au moment de l'ovulation. Elle se caractérise par une régénération de la couche fonctionnelle due à la migration et à la prolifération des cellules épithéliales des glandes de la couche basale vers la surface sous l'influence des œstrogènes (principalement l'œstradiol). Dans l'endomètre, de nouvelles glandes utérines se forment et des artères spirales se développent à partir de la couche basale. La durée de la phase peut varier. La température basale du corps est normale.
Phase de sécrétion- seconde moitié - dure de l'ovulation au début des règles. Les cellules épithéliales arrêtent de se diviser et s'hypertrophient. Les glandes utérines se dilatent et les cellules glandulaires sécrètent activement du glycogène, des glycoprotéines, des lipides et de la mucine. Dans les parties superficielles de la couche fonctionnelle, le nombre de cellules du tissu conjonctif augmente, autour desquelles se forment du collagène et des fibres réticulaires. Artères spirales deviennent plus alambiquées, se rapprochant de la surface de la membrane muqueuse. Si l'implantation de l'ovule fécondé n'a pas eu lieu, une diminution de la teneur en hormones stéroïdes ovariennes entraîne une torsion, une sclérose et une diminution de la lumière des artères spirales alimentant les deux tiers supérieurs de la couche fonctionnelle de l'endomètre. Il en résulte une détérioration du flux sanguin dans la couche fonctionnelle, une ischémie et un rejet, c'est-à-dire saignement.
Phase menstruelle- rejet de la couche fonctionnelle de l'endomètre, dure 3 à 7 jours.
Afin de déterminer le moment de l'ovulation, diverses méthodes de diagnostic fonctionnel des phases du cycle menstruel sont utilisées. Les paramètres suivants sont définis.
1. Température basale. Elle est associée à l'effet thermogénique de la progestérone (Fig. 10).
Jour de cycle 1 23456789 1010 11 12 13 14 15 16 17 1819 20 22 23 24 25 2627 2829
 Riz. dix. Température basale (rectale)
Riz. dix. Température basale (rectale)
2. Extensibilité de la glaire cervicale. Sous l'influence des œstrogènes, l'extensibilité du mucus augmente considérablement. Atteint des valeurs maximales pendant l'ovulation (Fig. 11)
3. L’effet de l’arborisation de la glaire cervicale (le phénomène « fougère »). Ce phénomène est plus prononcé pendant la période d'ovulation en raison de
une concentration élevée de sels de sodium qui précipitent en cristaux (symptôme de cristallisation), ressemblant extérieurement à une surface en forme d'arbre ou de fougère (Fig. 12).
4. Indice caryopycnotique - KPI (utilisant l'analyse microscopique d'un frottis vaginal).
 Riz. onze. Le degré d'extensibilité de la glaire cervicale
Riz. onze. Le degré d'extensibilité de la glaire cervicale
 Riz. 12. Le phénomène « fougère »
Riz. 12. Le phénomène « fougère »
L'IPC est le rapport entre les cellules kératinisées à noyaux pycnotiques (en pointillés) et l'ensemble des cellules de l'épithélium vaginal du frottis (Fig. 13, voir encadré). La valeur CPI la plus élevée correspond à la période d'ovulation - 70-80%, les jours restants du cycle menstruel - jusqu'à 30-40%.
Embryologie- la science de l'embryon, les lois de son développement. L'embryologie médicale étudie les schémas de développement de l'embryon humain, les caractéristiques structurelles, métaboliques et fonctionnelles de la barrière placentaire (système mère-placenta-fœtus), les causes des déformations et autres écarts par rapport à la norme, ainsi que les mécanismes de régulation. de l'embryogenèse.
Le concept d'embryogenèse comprend la période allant du moment de la fécondation à la naissance (pour les animaux vivipares), à l'éclosion des œufs (pour les animaux ovipares) et à la fin de la métamorphose (pour les animaux au stade de développement larvaire).
FERTILISATION
Transport de gamètes. Chez l'homme, le volume normal de l'éjaculat est d'environ 3 ml ; il contient en moyenne 350 millions de spermatozoïdes. Pour assurer la fécondation, le nombre total de spermatozoïdes doit être d'au moins 150 millions et leur concentration dans 1 ml doit être d'au moins 60 millions. En raison de leur grande mobilité, les spermatozoïdes dans des conditions optimales peuvent atteindre la cavité utérine en 30 minutes à 1 heure. et en 1,5 à 2 h situé dans la partie distale (ampullaire) de la trompe de Fallope, où se produit la fécondation. Les spermatozoïdes conservent leur capacité fécondante jusqu’à 2 jours.
L'ovocyte de premier ordre libéré par l'ovaire pendant l'ovulation a un diamètre d'environ 130 microns et est entouré d'une zone pellucide dense, ou membrane, et d'une couronne de cellules folliculaires, dont le nombre atteint 3 à 4 000. Il est prélevé. par les fimbriae de la trompe de Fallope (oviducte) et se déplace le long de celle-ci. C’est là que se termine la maturation de la cellule germinale. Dans ce cas, à la suite de la deuxième division, un ovocyte (œuf) de second ordre se forme, qui perd ses centrioles et ainsi la capacité de se diviser. Le noyau d'un œuf humain contient 23 chromosomes ; l’un d’eux est le chromosome sexuel X.
L’ovule humain épuise généralement ses réserves de nutriments dans les 12 à 24 heures suivant l’ovulation, puis meurt s’il n’est pas fécondé.
Fertilisation se produit dans la partie ampullaire de l'oviducte. Conditions optimales pour l'interaction des spermatozoïdes avec l'ovule
généralement créé dans les 12 heures après l’ovulation. Lors de l'insémination, de nombreux spermatozoïdes s'approchent de l'ovule et entrent en contact avec sa membrane. L'œuf commence à effectuer des mouvements de rotation autour de son axe à une vitesse de 4 rotations par minute. Ces mouvements sont provoqués par le battement des flagelles des spermatozoïdes et durent environ 12 heures. Lors de l'interaction des cellules germinales mâles et femelles, un certain nombre de changements se produisent. Les spermatozoïdes sont caractérisés par des phénomènes de capacitation et de réaction acrosomale. La capacitation est le processus d'activation des spermatozoïdes dans la trompe de Fallope sous l'influence de la sécrétion muqueuse des cellules glandulaires. La progestérone active la sécrétion des cellules glandulaires. Après la capacitation, une réaction acrosomale s'ensuit, au cours de laquelle les enzymes hyaluronidase et trypsine sont libérées du sperme. La hyaluronidase décompose l'acide hyaluronique contenu dans la zone pellucide. La trypsine décompose les protéines du cytolemme des cellules de l'œuf et de la couronne radiée. En conséquence, les cellules de la couronne radiée se dissocient et la zone pellucide se dissout.
Dans l'ovule, le cytolemme dans la zone de fixation du sperme forme un tubercule de levage dans lequel pénètre un spermatozoïde, et une membrane dense apparaît - la membrane de fécondation, qui empêche l'entrée d'autres spermatozoïdes et le phénomène de polyspermie. Les noyaux des cellules germinales femelles et mâles se transforment en pronoyaux, se rapprochent et le stade syncaryon commence. Se pose zygote, et à la fin du 1er jour après le début de la fécondation Se séparer.
Le sexe de l'enfant dépend des chromosomes sexuels du père. En raison de la plus grande sensibilité des embryons mâles aux effets néfastes de divers facteurs, le nombre de nouveau-nés garçons est inférieur à celui des filles : pour 100 garçons, 105 filles naissent.
Le mouvement de l'ovule fécondé est assuré par des contractions péristaltiques des muscles du tube et le scintillement des cils de l'épithélium. L'embryon se nourrit de petites réserves de jaune dans l'œuf et, éventuellement, du contenu de la trompe de Fallope.
Le transport de l'embryon vers l'utérus s'effectue dans un environnement immunosuppresseur, dans lequel se forment les spermatozoïdes, le liquide du blastocyste, UN Protéine 2-utérine (commence à être produite par l'épithélium glandulaire de l'endomètre dans les jours qui suivent l'ovulation) et facteur de grossesse précoce (EGF), décrit pour la première fois par H. Morton en 1974. L'EGF est produit par l'ovule 46 à 48 heures après
le fécondation et est l’un des premiers indicateurs de grossesse et le premier agent immunosuppresseur qui prévient le rejet des blastocystes. Facteurs de protection immunologique :
UN 2 - protéine de la glande endomètre ;
Facteur de grossesse précoce de l'œuf ;
Protéines immunobloquantes des syncytiotrophoblastes ;
HCG et lactogène placentaire (PL) ;
Lycoprotéines fibrinoïdes placentaires ;
Propriétés protéolytiques du trophoblaste.
Fragmentation de l'embryon humain commence à la fin du 1er jour et se poursuit pendant 3 à 4 jours après la fécondation (à mesure que l'embryon se déplace vers l'utérus). La fragmentation du zygote humain est complète, inégale, asynchrone. Le premier jour, cela se produit lentement. La première division est terminée après 30 heures ; dans ce cas, le sillon de clivage longe le méridien et deux blastomères se forment. Le stade à deux blastomères est suivi du stade à quatre blastomères. Après 40 heures, quatre cellules se forment (Fig. 14, voir encadré).
Dès les premières divisions, deux types de blastomères se forment : « foncés » et « clairs ». Les blastomères « clairs » se fragmentent plus rapidement et sont situés en une seule couche autour des « sombres », qui se retrouvent au milieu de l'embryon. À partir des blastomères « légers » de surface, un trophoblaste naît ensuite, reliant l'embryon à l'organisme maternel et lui fournissant de la nutrition. Les blastomères internes « sombres » forment l'embryoblaste - à partir de lui, le corps de l'embryon et tous les autres organes extra-embryonnaires, à l'exception du trophoblaste, sont formés. Au moment où le blastocyste pénètre dans l'utérus, sa taille augmente en raison d'une augmentation du nombre de blastomères et du volume de liquide due à l'absorption accrue des sécrétions des glandes utérines par le trophoblaste et à la production active de liquide par le trophoblaste lui-même.
Dans le trophoblaste, le nombre de lysosomes augmente, dans lesquels s'accumulent des enzymes qui assurent la lyse du tissu utérin et facilitent ainsi l'introduction de l'embryon dans l'épaisseur de la muqueuse utérine, c'est-à-dire nidation. L'implantation (nidation) débute le 7ème jour après la fécondation et dure environ 40 heures (Fig. 15, voir encadré). Dans ce cas, le blastocyste est complètement entouré de tissu endométrial - la caduque.
La couche trophoblastique se différencie bientôt en la couche externe - le syncytiotrophoblaste, qui est constamment reconstituée en noyaux et en cytoplasme en raison de la couche interne sous-jacente du cytotrophoblaste (couche de Langhans), puisque la division nucléaire n'est observée que dans le cytotrophoblaste. Le troisième dérivé du trophoblaste est non prolifératif et est un type de cellule mononucléaire initialement appelé « cellules X » et également connu sous le nom de « trophoblaste intermédiaire ». Il s'agit du principal type de cellules qui composent la plate-forme placentaire et, avec les cellules de la caduque, pénètrent dans les artères spirales maternelles et forment également la majeure partie des cellules des cloisons placentaires. Les cellules X sont la principale source de lactogène placentaire humain (HPL - lactogène placentaire humain) et de grandes quantités de protéines essentielles à la grossesse (MBP - protéine basique majeure)
Durant les 2 premières semaines, le trophoblaste consomme les produits de dégradation des tissus maternels (alimentation de type histiotrophique). Ensuite, le syncytiotrophoblaste, se développant sous forme de villosités et produisant des enzymes protéolytiques, pénètre dans l'utérus, détruit les vaisseaux déciduaux maternels, permettant ainsi au sang maternel de s'écouler dans des lacunes inégales - qui sont le futur « espace intervilleux ». Ainsi, le trophoblaste entre en contact direct avec le sang des vaisseaux maternels et l'embryon commence à se nourrir directement du sang maternel (alimentation de type hématotrophe). La circulation sanguine complète chez le fœtus s'établit environ au cours de la 5ème semaine après la fécondation.
Une fois l’implantation terminée, commence une période très importante d’organogenèse et de placentation dans le développement de l’embryon. Du 20e au 21e jour, le corps de l'embryon se sépare des organes extra-embryonnaires et se produit la formation finale des primordiums axiaux. L'organogenèse est terminée entre la 12e et la 16e semaine de la vie intra-utérine.
Les périodes de développement prénatal sont indiquées sur la Fig. 16.
La masse embryonnaire se différencie, les feuillets germinaux se forment : 1) ectoderme ; 2) mésoderme ; 3) endoderme. Ils sont également différenciés (Fig. 17, voir encadré).
Le tube neural est formé à partir de l'ectoderme. La fermeture du tube neural commence dans la région cervicale, puis s'étend vers l'arrière et le crâne, là où se forment les vésicules médullaires. Vers le 25ème jour, le tube neural est complètement fermé et du milieu extérieur
 Riz. 16. Périodes de développement prénatal
Riz. 16. Périodes de développement prénatal
Seules deux ouvertures non fermées communiquent entre elles aux extrémités antérieure et postérieure : les neuropores antérieur et postérieur. Après 5 à 6 jours supplémentaires, les deux neuropores sont envahis. Lorsque les parois latérales des plis neuraux se ferment et que le tube neural se forme, ce qu'on appelle la crête neurale apparaît. Les cellules de la crête neurale sont capables de migrer. Dans le tronc, les cellules migratrices forment les ganglions parasympathiques et sympathiques et la médullosurrénale. Certaines cellules restent dans la région de la crête neurale, elles sont segmentées et donnent naissance aux ganglions spinaux.
Différenciation mésoderme commence le 20ème jour de l'embryogenèse.
Les cellules du mésoderme se précipitent vers la surface interne de la cavité du blastocyste et se différencient en tissu conjonctif du chorion et des villosités. L'endroit où ces cellules quittent l'embryon devient le cordon ombilical, dans lequel se développent les vaisseaux allantoïdiens du futur placenta.
Les changements dans l'embryon lui-même s'expriment par le fait que les sections dorsales des feuilles mésodermiques sont divisées en segments denses situés sur les côtés de la notocorde - les somites. Le processus de formation de segments, ou somites, commence dans la partie tête de l'embryon et s'étend dans la direction caudale. Et si au 22ème jour de développement l'embryon a 7 paires de segments, alors au 35ème jour il y en a 44 paires. Dans le processus de différenciation du mésoderme, un rudiment néphrogénique et un embryon
Le premier rudiment du tissu conjonctif est le mésenchyme. Les cellules ecto- et endodermiques participent partiellement à la formation du mésenchyme.
Endoderme forme une cavité - l'intestin primaire, le futur tube digestif, qui se développe tout au long de l'étape de formation du sac vitellin. La séparation de l'endoderme intestinal commence par l'apparition du pli du tronc qui, en s'enfonçant plus profondément, sépare l'endoderme embryonnaire - l'intestin primaire - de l'endoderme extra-embryonnaire - le sac vitellin. Au début de la 4ème semaine, une invagination ectodermique se forme à l'extrémité antérieure de l'embryon - la fosse buccale. En s'approfondissant, la fosse atteint l'extrémité antérieure de l'intestin et, après avoir traversé la membrane qui les sépare, elle se transforme en ouverture buccale de l'enfant à naître.
Le sac vitellin et le tube digestif restent reliés pendant un certain temps par le canal omphalomésentérique (tige vitellue), se terminant par un potentiel diverticule de Meckel. La tige vitelline, comme le sac vitellin, s'atrophie par la suite.
Ainsi, le sac vitellin, formé par l'endoderme extraembryonnaire et le mésoderme extraembryonnaire, participe activement à la nutrition et à la respiration de l'embryon humain pendant une très courte période. Le rôle principal du sac vitellin est hématopoïétique. En tant qu'organe hématopoïétique, il fonctionne jusqu'à la 7-8ème semaine, puis subit un développement inverse. Dans la paroi du sac vitellin, des cellules germinales primaires - des gonoblastes - se forment, migrant de celle-ci avec le sang vers les rudiments des gonades.
Dans la partie postérieure de l'embryon, l'intestin résultant comprend également la partie de l'endoderme d'où provient l'excroissance endodermique de l'allantoïde.
L'allantoïde est un petit processus en forme de doigt de l'endoderme qui se développe en tige amniotique. Chez l'homme, l'allantoïde n'est pas très développé, mais son importance pour assurer la nutrition et la respiration de l'embryon est encore grande, puisque des vaisseaux se développent le long de lui vers le chorion dont les dernières branches se trouvent dans le stroma des villosités. Au 2ème mois de l'embryogenèse, l'allantoïde est réduite.
En figue. La figure 18 (voir encadré) montre à quoi ressemble un embryon à 4-5 semaines.
Pendant les périodes d'organogenèse (Fig. 19) et de placentation, en raison de l'action pathogène de facteurs environnementaux sur l'embryon et le fœtus, les organes et systèmes qui se trouvent dans cet état sont principalement affectés.
temps en cours de différenciation. Pour divers ébauches d'organes embryonnaires, les périodes critiques ne coïncident pas dans le temps. Par conséquent, l’action d’un facteur dommageable provoque généralement des déformations de divers organes et systèmes. La phase de développement la plus sensible correspond aux 3 à 6 premières semaines de l'ontogenèse (la deuxième période critique du développement).
 Riz. 19. Périodes d'organogenèse
Riz. 19. Périodes d'organogenèse
CROISSANCE INTRA-utérine DU FŒTAL
La dynamique de la croissance fœtale dans l'utérus est assurée par l'interaction du potentiel génétique de chaque fœtus et de l'environnement intra-utérin, qui est principalement associé au fonctionnement du placenta et à l'homéostasie maternelle. La dynamique de la croissance fœtale au cours de la grossesse physiologique correspond à l'âge gestationnel (tableau 1).
Tableau 1
Dynamique de la croissance fœtale
Après 27 semaines de grossesse, la dynamique de croissance est influencée par le sexe du fœtus (Tableau 2).
Tableau 2a
Semaines de gestation | Centiles de masse pour les garçons, g (A.V. Mazurin, I.M. Vorontsov, 2000) |
||||
Tableau 2b
Dynamique de croissance fœtale selon le sexe
Semaines de gestation | Centiles de masse des filles, g (A.V. Mazurin, I.M. Vorontsov, 2000) |
||||
L'écart entre la taille du fœtus et la période réelle de grossesse est défini par la notion de « retard de croissance intra-utérin » (RCIU) du fœtus. Le critère international pour le RCIU est un poids et/ou une taille fœtale inférieurs à la normale pour l'âge gestationnel (10e centile et moins). Le syndrome IUGR est l'une des manifestations cliniques de l'insuffisance placentaire.
Placenta- un organe extra-embryonnaire par lequel s'établit une connexion entre l'embryon et le corps de la mère. Le placenta humain appartient au type de placenta villeux hémochorial discoïde. La formation du placenta commence à la 3ème semaine, lorsque les vaisseaux commencent à se développer en villosités secondaires (épithéliomésenchymateuses), formant des villosités tertiaires, et se termine à 14-16 semaines de grossesse.
Le placenta est divisé en partie embryonnaire, ou fœtale, et en partie maternelle, ou utérine.
La partie fœtale est représentée par un chorion ramifié et la membrane amniotique qui y est attachée, et la partie maternelle est représentée par une partie basale modifiée de l'endomètre.
La partie embryonnaire ou fœtale du placenta à la fin du 3ème mois est représentée par une plaque chorionique ramifiée, constituée de tissu conjonctif fibreux (collagène) recouvert de cyto- et syncytiotrophoblaste. Les villosités ramifiées du chorion (tige, ou ancre, villosités) ne sont bien développées que du côté faisant face
myomètre. Ici, ils traversent toute l'épaisseur du placenta et, avec leurs sommets, sont immergés dans la partie basale de l'endomètre détruit.
L'unité structurelle et fonctionnelle du placenta formé est le cotylédon, formé par les villosités souches et ses branches secondaires et tertiaires. Le nombre total de cotylédons dans le placenta atteint 200.
La partie maternelle du placenta est représentée par la plaque basale et les septa de tissu conjonctif qui séparent les cotylédons les uns des autres, ainsi que par des lacunes remplies de sang maternel.
À la surface de la plaque basale, face aux villosités choriales, se trouve une substance amorphe - le fibrinoïde de Rohr. Les cellules trophoblastiques de la lame basale, ainsi que les fibrinoïdes, jouent un rôle important dans l'assurance de l'homéostasie immunologique dans le système mère-fœtus.
Le sang dans les lacunes se renouvelle continuellement. Il provient des artères utérines, qui entrent ici depuis la muqueuse musculaire de l'utérus. Ces artères longent les cloisons placentaires et débouchent sur des lacunes. Le sang maternel coule du placenta par des veines provenant des lacunes.
Le sang de la mère et celui du fœtus circulent dans des systèmes vasculaires indépendants et ne se mélangent pas. La barrière hémochorionique séparant les deux flux sanguins est constituée de l'endothélium des vaisseaux fœtaux, des vaisseaux du tissu conjonctif environnants, de l'épithélium des villosités choriales (cytotrophoblaste, syncytiotrophoblaste) et, en outre, du fibrinoïde, qui recouvre à certains endroits les villosités de l'extérieur. .
Le placenta remplit des fonctions trophiques, excrétrices (pour le fœtus), endocriniennes (produit de l'hCG, de la progestérone, du PL, des œstrogènes, etc.), protectrices (y compris la protection immunologique).
Valeur HCG
Stimule la production de progestérone par le corps jaune.
Stimule les cellules de Leydig des fœtus mâles et la production de testostérone.
Détermine le développement des organes génitaux masculins.
C'est un marqueur précoce de grossesse.
C'est un critère d'évaluation de l'efficacité du traitement des tumeurs trophoblastiques, ainsi qu'un inducteur d'ovulation en raison de sa similitude biologique avec l'HTP.
Propriétés PL
Participe à la défense immunologique - inhibe les lymphocytes maternels.
Stimule la lipolyse et augmente la concentration en acides gras libres.
Inhibe la gluconéogenèse maternelle.
Augmente les niveaux d'insuline plasmatique.
Stimule la synthèse des protéines et des acides aminés grâce à l'effet insulinogène.
La concentration de PL dépend du poids du placenta.
Membrane amniotique. Il est avasculaire et forme la paroi la plus interne du réceptacle du fruit. Sa fonction principale est la production de liquide amniotique, qui fournit un environnement au corps en développement et le protège des dommages mécaniques. L'épithélium de l'amnios, face à sa cavité, sécrète du liquide amniotique et participe également à leur réabsorption. Dans ce cas, dans l'épithélium de l'amnios recouvrant le disque placentaire, la sécrétion se produit principalement, et dans l'épithélium de l'amnios extraplacentaire, la résorption du liquide amniotique se produit principalement. Le liquide amniotique crée l'environnement aqueux nécessaire au développement de l'embryon, en maintenant la composition et la concentration de sels nécessaires dans le liquide amniotique jusqu'à la fin de la grossesse. L'amnios remplit également une fonction protectrice en empêchant les agents nocifs de pénétrer dans le fœtus.
L'amnios est vaguement relié au chorion, dans lequel se trouvent les vaisseaux fœtaux. Son attachement au chorion intervient vers la 12ème semaine de grossesse ; Avant cela, il y a un espace rempli de liquide entre l'amnios et le chorion. De plus, l’amnios bouge souvent pendant la grossesse et peut même se détacher bien avant la naissance. Il forme aussi parfois des cordons qui, s'ils entrent en contact avec le fœtus, peuvent provoquer des amputations prénatales et d'autres déformations. L'amnios étant relié au cordon ombilical et y étant étroitement attaché, les restes des cordons se retrouvent le plus souvent au site de fixation du cordon ombilical.
Cordon ombilical
Le cordon ombilical est formé principalement de mésenchyme situé dans la tige amniotique et la tige vitelline. L'allantoïde et les vaisseaux qui y poussent participent également à la formation de la moelle. En surface, toutes ces formations sont entourées par la membrane amniotique. La tige vitelline et l'allantoïde sont rapidement réduits et seuls leurs restes se retrouvent dans le cordon ombilical du nouveau-né.
Le cordon ombilical formé est une formation de tissu conjonctif élastique dans laquelle passent deux artères ombilicales et une veine ombilicale. Il est formé de tissu gélatineux (muqueux) typique, qui contient une énorme quantité d'acide hyaluronique. C'est ce tissu, appelé gelée de Wharton, qui assure la turgescence et l'élasticité du cordon. Il protège les vaisseaux ombilicaux de la compression, assurant ainsi un apport continu de nutriments et d'oxygène à l'embryon.
Normalement, le cordon ombilical est attaché au disque placentaire (attache centrale), dans 7% il y a une attache marginale (battledor) et à 1% - sur les membranes (fixation mécanique). Les attachements anormaux sont plus fréquents dans les grossesses multiples. L'implantation du placenta n'est pas associée à des anomalies fœtales, mais peut être dangereuse en raison de l'incidence accrue de thrombose vasculaire et de la possibilité de saignement provenant de vaisseaux rompus lors de l'accouchement.
La longueur du cordon ombilical est largement déterminée par l'activité motrice du fœtus. Ainsi, un cordon ombilical court indique souvent son immobilité due à une pathologie neuromusculaire ou à des fusions amniotiques. Au contraire, un cordon ombilical long est parfois le résultat d'une activité motrice accrue du fœtus.
Une seule artère du cordon ombilical survient dans plus de 1 % des cas, plus souvent en cas de grossesses multiples. Environ la moitié de ces nouveau-nés présentent des anomalies congénitales, dont certaines devraient être activement diagnostiquées, ainsi que d'autres problèmes périnatals. Cependant, une seule artère du cordon ombilical peut être présente chez un nouveau-né tout à fait normal ; alors cette découverte signale seulement la nécessité d’être prudent quant à la présence d’une pathologie chez ce nouveau-né.
Malgré le fait que le corps de la mère et le fœtus soient génétiquement étrangers dans la composition des protéines, un conflit immunologique ne se produit généralement pas.
des promenades. Ceci est assuré par un certain nombre de facteurs ; Parmi ceux-ci, les suivants sont particulièrement importants :
1 - protéines synthétisées par les syncytiotrophoblastes qui inhibent la réponse immunitaire du corps maternel ;
2 - hCG et PL, localisés en concentrations élevées à la surface du syncytiotrophoblaste, participant à l'inhibition des lymphocytes maternels ;
3 - effet immunomasquant des glycoprotéines du fibrinoïde placentaire, chargées, comme les lymphocytes de lavage du sang, négativement ;
4 - propriétés protéolytiques du trophoblaste, qui contribuent également à l'inactivation des protéines étrangères ;
5 - liquide amniotique contenant des anticorps qui bloquent les antigènes A et B (contenus dans le sang d'une femme enceinte) et les empêchent de pénétrer dans le sang du fœtus en cas de grossesse incompatible.
Dans le processus de formation du système mère-fœtus, il existe un certain nombre de périodes critiques qui sont les plus importantes pour établir une interaction entre les deux systèmes et pour créer des conditions optimales pour le développement du fœtus.
Dans l'ontogenèse humaine, plusieurs périodes critiques du développement peuvent être distinguées : la progenèse, l'embryogenèse et la vie postnatale. Ceux-ci inclus:
1) développement des cellules germinales - oogenèse et spermatogenèse ;
2) la fécondation ;
3) implantation (7-8 jours d'embryogenèse) ;
4) développement des ébauches des organes axiaux et formation du placenta (3-8ème semaine de développement) ;
5) stade de croissance cérébrale accrue (15-20e semaine) ;
6) formation des principaux systèmes fonctionnels du corps et différenciation de l'appareil reproducteur (20-24e semaine) ;
7) naissance ;
8) période néonatale (jusqu'à 1 an) ;
9) puberté (11-16 ans).